Si tratta di un tessuto altamente vascolarizzato e povero di connettivo (che si ritrova solo nel SNP, mentre nel SNC accompagna solo i vasi); si compone di cellule nervose propriamente dette (i neuroni) e di cellule della glia (a funzione di sostegno, di protezione e trofica); i neuroni sono cellule perenni caratterizzate da un corpo cellulare (o soma o pirenoforo) dal quale si dipartono più o meno prolungamenti, di cui uno detto
assone o neurite, a carattere di trasporto centrifugo dell'impulso nervoso, solitamente più lungo e ramificato distalmente, e gli altri detti
dendriti, a carattere ricettivo, con ramificazioni più prossimali e spine dendritiche. Il neurone è infatti eccitabile, capace di generare un impulso a seguito di uno stimolo e di condurlo trasmettendolo alle altre cellule nervose attraverso sinapsi.
Data la loro elevata attività, i neuroni presentano un nucleo eucromatico, vescicoloso, presentante uno o due nucleoli (a causa dell'elevata trascrizione e produzione di ribosomi); inoltre, sempre per la stessa ragione, il citoplasma del corpo cellulare e quello dendritico è ricco di RER, che se trattato con coloranti particolari si presenta a zolle (zolle di Nissl); l'assone, che si diparte da una zona della cellula detta cono di emergenza, non presenta gli stessi organuli del resto del neurone, ma microtubuli per il trasporto di organuli e sostanze alle estremità sinaptiche (dove è necessaria molta energia e quindi la presenza di mitocondri).
I neuroni si classificano in base alla forma in neuroni
"solo dendriti" con prolungamenti corti di cui non si distingue l'assone, neuroni
unipolari aventi solo l'assone, neuroni
bipolari con un solo dendrite e l'assone che emergono da punti diametralmente opposti del soma, neuroni
pseudounipolari a T con un solo lungo dendrite e l'assone che si dipartono entrambi da un tratto comune, e neuroni
multipolari con molto dendriti corti e l'assone.
Essendo i neuroni cellule perenni, che se degradate non si rigenerano, esse sono protette dalle cellule della glia che ne avvolgono sia i corpi cellulari che i prolungamenti; le cellule gliali svolgono però anche altre funzioni; in particolare nel SNC ritroviamo
astrociti (che contribuiscono a formare la barriera ematoencefalica),
oligodendrociti (che emettono prolungamenti citoplasmatici i quali avvolgono tratti di più assoni proteggendoli),
cellule ependimali (a carattere epiteliale, rivestono il canale ependimale e i ventricoli cerebrali producendo liquido cefalorachidiano) e
cellule della microglia (che fanno parte del sistema dei monociti e macrofagi ed eliminano i detriti); nel SNP ritroviamo
cellule di Schwann (che possono avvolgere un tratto di assone col loro corpo cellulare formando fibre mieliniche oppure contenere tratti di diversi assoni in tasche citoplasmatiche formando fibre amieliniche) e
cellule satelliti (che abbracciano il soma dei neuroni ad esempio pseudounipolari nei gangli spinali).
Le fibre mieliniche sono quindi assoni sui quali cellule di Schwann o oligodendrociti hanno avvolto una porzione della loro membrana svuotata dal citoplasma (mielina), così da isolare elettricamente il prolungamento e da permettere una trasmissione più rapida dell'impulso, il quale salta tra i brevi tratti scoperti da mielina e ricoperti solo dai corpi delle cellule gliali; le fibre amieliniche, non essendo assoni isolati da avvolgimenti di sola membrana, ma protetti dal citoplasma delle cellule gliali, trasmettono più lentamente l'impulso.
#NEURONI
Si osservano principalmente due strutture con la stessa morfologia; in quella sottostante, meglio visibile, si osserva una zona a macchie di leopardo (citoplasma con zolle di Nissl), da cui sembrano dipartirsi prolungamenti (dendriti e assone); vi è una zona tondeggiante più chiara e vacuolosa al suo interno, il nucleo, contenente un'altra struttura tondeggiante, il nucleolo. Si tratta quindi di neuroni, circondati da nuclei di altre cellule, probabilmente gliali.
In questo preparato si riescono a vedere bene i profili di cellule nervose con il loro nucleo contenente il nucleolo, e con i loro prolungamenti più o meno definiti.
#NERVO
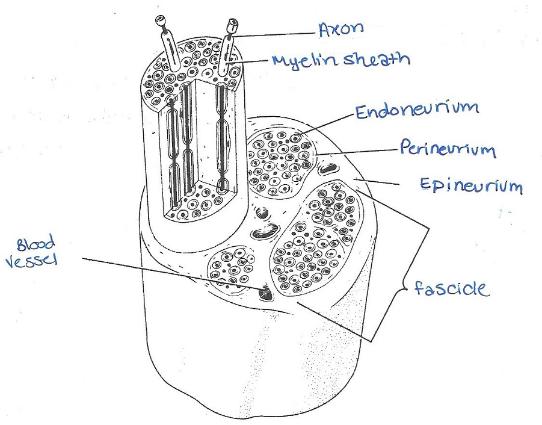
I nervi fanno parte del SNP e sono costituiti da fasci paralleli di fibre assoniche (assoni circondati da mielina) raggruppati e organizzati con del connettivo in modo molto simile a quelle muscolari scheletriche (endonevrio, perinevrio ed epinevrio)
Si osservano strutture tondeggianti strettamente impacchettate in strutture più grandi, avvolte da connettivo apparentemente denso (azzurro, epinevrio); all'interno delle piccole unità tondeggianti otticamente vuote si osserva un puntino centrale più scuro; si tratta dell'assone che era circondato da mielina, la quale essendo formata da lipidi (membrana delle cellule di Schwann avvolta attorno al prolungamento) è andata persa con le tecniche di fissazione chimica; questa è quindi una sezione trasversale di un nervo, in cui si osservano le fibre mieliniche immerse nell'endonevrio e organizzate dal perinevrio in fasci; più fasci racchiusi nell'epinevrio formano il nervo.
#FIBRE MIELINICHE
Si osservano strutture "cave" dello stesso genere disposte parallelamente e con un asse longitudinale prevalente; si tratta di fibre mieliniche, fissate con congelamento e trattate con tetrossido d'osmio; sono ben visibili i nodi di Ranvier (zone dell'assone dove la mielina è assente e che permettono il passaggio dell'impulso nervoso); poco visibili in questo preparato sono solitamente presenti particolari formazioni dette incisure dello Schmidt-Lanterman, dovute alla presenza di piccole quantità di citoplasma delle cellule di Schwann tra gli strati di mielina che appaiono come incisioni interne dello strato isolante.
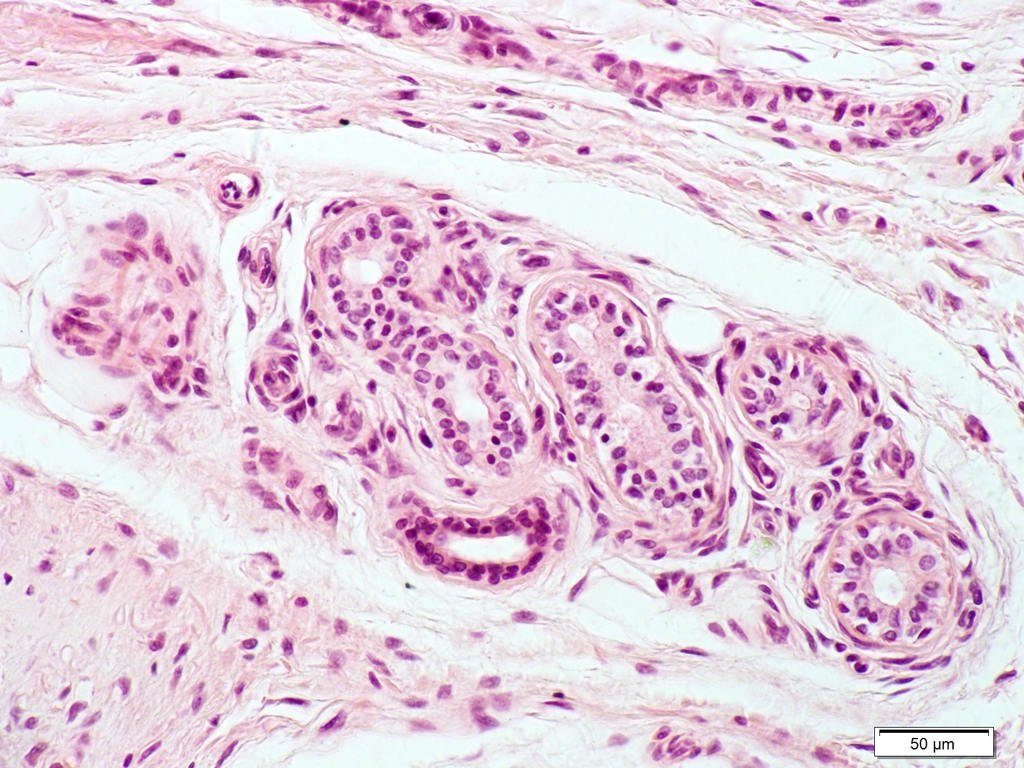
#GANGLIO SPINALE è una struttura del SNP, di forma ellissoidale, rivestito da una capsula connettivale e avente uno stroma di connettivo reticolare in cui sono immersi i pirenofori e il primo tratto dei prolungamenti di neuroni pseudounipolari sensoriali, circondati da cellule gliali
Si osservano cellule dal citoplasma eterogeneo, con un nucleo vescicoloso con uno o due nucleoli; si tratta di neuroni, coi corpi cellulari abbracciati da un numero molto elevato di cellule (cellule satelliti) e circondati da altri nuclei di cellule ipoteticamente di Schwann e connettivali; i neuroni sono immersi in un connettivo lasso; si tratta di un ganglio spinale e di neuroni pseudounipolari; se fosse possibile avere un ingrandimento più basso del preparato si potrebbe distinguere anche la capsula di connettivo più denso.






































.jpg)