In tale tessuto la contrazione è basata sul lavoro compiuto da particolari fibre proteiche:
- filamenti di actina, costituiti da monomeri di actina globulare associati a formare una doppia catena spiralizzata;
 - filamenti di miosina, costituiti da molecole di miosina associate in modo sfalsato a formare due fasci uniti per le code; sotto le teste della miosina è presente una zona detta a cerniera che è un punto di snodo e ne permette la flessione; la contrazione avviene perchè le teste della miosina si agganciano all'actina e si flettono facendo scorrere i due filamenti l'uno sull'altro; attraverso quindi una serie di attacchi-flessioni-stacchi delle teste di miosina si ha un'accorciamento della fibra;
- filamenti di miosina, costituiti da molecole di miosina associate in modo sfalsato a formare due fasci uniti per le code; sotto le teste della miosina è presente una zona detta a cerniera che è un punto di snodo e ne permette la flessione; la contrazione avviene perchè le teste della miosina si agganciano all'actina e si flettono facendo scorrere i due filamenti l'uno sull'altro; attraverso quindi una serie di attacchi-flessioni-stacchi delle teste di miosina si ha un'accorciamento della fibra;- tropomiosina, una proteina che accompagna tutto il filamento di actina coprendone i siti di legame per le teste della miosina, impedendo così la contrazione quando non è necessaria;
- troponina, composta da tre proteine globulari di cui una (subunità I) lega l'actina, una (subunità T) lega la tropomiosina tenendola in sede, e l'altra (subunità C) può legare ioni calcio provocando lo scivolamento della tropomiosina e quindi la messa in evidenza dei siti di legame dell'actina per la miosina.

Tali filamenti sono organizzati in strutture dette sarcomeri, i quali sono strutturati con due linee Z parallele alle estremità, costituite da proteine, le quali hanno agganciati perpendicolarmente filamenti di actina che si portano centralmente alla struttura formando una sorta di registro; tra ogni coppia di filamenti di actina, centralmente e collegati da proteine (disposte su una linea M) sono disposti i filamenti pesanti di miosina; queste strutture che sostituiscono le unità fondamentali delle miofibrille, sono anche le responsabili delle bande nella muscolatura striata: le larghe bande scure centrali al sarcomero (bande A) dovute alla miosina, e le bande chiare tagliate a metà dalle linee Z (bande I) dovute i filamenti di actina.


Inoltre nella fibra muscolare va considerato un altro tipo di organizzazione, e cioè quello responsabile del rilascio degli ioni calcio (accumulati nel REL della fibra), e quindi della contrazione; infatti le miofibrille occupano quasi completamente il sarcoplasma della fibra, e tra esse decorre il REL che le avvolge con le sue cisterne disposte sia parallelamente che perpendicolarmente ad esse, con una morfologia diversa nei diversi punti del sarcomero. In corrispondenza del margine tra la semibanda I e la banda A vi sono strutture particolari dette triadi, costituite da due cisterne terminali del REL tra le quali è presente un'invaginazione della membrana plasmatica della cellula (tubulo T); le cisterne terminali delle triadi adiacenti sono connesse da elementi longitudinali che a livello delle linee M (o delle bande H, che dir si voglia) si fondono a formare una rete di cisterne (cisterna fenestrata).

Essendo la contrazione muscolare un processo energeticamente molto dispendioso, tra le miofibrille sono presenti anche molti mitocondri; al termine della depolarizzazione gli ioni calcio vengono nuovamente raccolti nelle cisterne del REL.
Tessuto cardiaco
La fibra muscolare cardiaca non possiede un'organizzazione così ben definita come quella scheletrica, ma il principio della contrazione è identico; l'unica differenza nell'organizzazione è che nel miocardio le cisterne terminali del REL sono discontinue e quindi non si parla di triadi ma di diadi con i tubuli T, i quali non sono posizionati tra la semibanda I e la banda A ma sono in corrispondenza della linea Z; si ha così una diade per ogni linea Z delle miofibrille.

Tessuto liscio
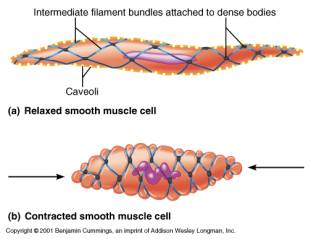
Le fibre per la contrazione, il REL e le invaginazioni di membrana non sono organizzati come nel tessuto striato bensì la cellula presenta invaginazioni in direzioni diverse dette caveole (equivalenti dei tubuli T) e subito al di sotto della membrana plasmatica sono presenti dei corpi densi per l'aggancio delle fibre di actina (come le linee Z); le molecole di miosina sono presenti ripiegate su loro stesse e solo al momento della contrazione si associano alle altre e alle fibre di actina a formare strutture dall'andamento obliquo rispetto all'asse longitudinale della cellula. In questo modo la contrazione provoca un avvitamento della fibra muscolare.
Nessun commento:
Posta un commento